

導入:細胞は「壊される」のを待っているわけではない
放射線がDNAに傷をつけた後、細胞には過酷な選択が突きつけられる。「完璧に直して生き残るか」「直せずに死ぬか」、あるいは「中途半端に直してがん化するか」である。
放射線の影響は、細胞の種類や「今、何をしている時間か(細胞周期)」によって劇的に変化する。本稿では、放射線感受性の絶対ルールである「ベルゴリー・トリボンドーの法則」から、命運を分けるDNA修復のメカニズムまでを、論理的に解明していく。
第1章:放射線感受性の支配ルール(B-T法則と幹細胞)
どの細胞が放射線に弱いのか。その答えは100年以上前に提唱された一つの法則に集約されている。
1-1. ベルゴリー・トリボンドー(B-T)の法則
放射線感受性の高さを決定する3つの条件である。これを一言で言えば、**「未来への可能性に満ち、忙しく働いている細胞ほど弱い」**ということである。
- 分裂能力が高い細胞(分裂頻度が高い) 細胞分裂の回数が多いほど、DNAが剥き出しになる機会が増え、致命的なダメージを受けやすくなる。
- 将来の分裂回数が多い細胞 これから何度も分裂を繰り返す予定の細胞は、小さな傷が将来的に大きなエラーとして増幅されるため、感受性が高い。
- 未分化な細胞(特定の役割が決まっていない) 特定の臓器の細胞(分化細胞)になる前の、いわば「何にでもなれる卵」の状態の細胞は、構造が単純で放射線に弱い。
【結論:感受性が高い細胞の正体】 この3条件をすべて満たすのが、いわゆる**「幹細胞(Stem Cell)」**である。幹細胞は際限なく増殖し、未分化であるため、放射線に対して極めて高い感受性を持つ。
1-2. 感受性とアポトーシス
感受性が高い細胞は、放射線を受けた際に無理に生き延びようとせず、自ら死を選ぶ**「アポトーシス(プログラムされた細胞死)」**を起こしやすい。
これは個体全体を守るための防衛反応でもある。修復ミスを抱えたまま分裂して「がん」になるくらいなら、その場で消えてしまった方が安全だからである。そのため、感受性が高い組織ほど、照射直後に急速に細胞が減少するという特徴がある。
第2章:細胞周期による感受性の変化(タイミングの科学)
細胞は、分裂(M期)と準備(G1期・S期・G2期)というサイクル(細胞周期)を繰り返しており、放射線に対する「感受性(効き目)」はどの段階にいるかによって劇的に変化する。
感受性を前期後期など暗記するのは大変なので下記のようにサイクルを書いて高感受性の部分だけ下線を引く方式で暗記するといい。

2-1. 国試頻出:感受性が高い時期の「明文化」
放射線技師国家試験において、細胞周期と感受性の関係は極めて重要である。特に、感受性が高くなるタイミングの定義は定番の出題ポイントとなるため、以下の2点を正確に記憶すること。
- G2後半からM期終わりまで:細胞周期を通じて最も感受性が高い(放射線に弱い)。
- G1後期からS前期まで:再び感受性が高まる第2のピーク。
逆に、**最も感受性が低い(放射線に強い=抵抗性が高い)のは「後期S期」**である。
2-2. なぜその時期に弱く、あるいは強いのか?(ロジック解説)
この感受性の差には、DNAの形態や修復機能といった明確な理由が存在する。
- G2後半からM期終わり(高感受性)の理由: DNAがギュッと凝縮して染色体(M期)となり、細胞分裂が行われる。この時期は「細胞分裂」に全リソースを注いでおり、DNA修復酵素の働きが低下しているため、ダメージがそのまま「死」へ直結しやすい。
- G1後期からS前期(高感受性)の理由: DNAの複製を開始する準備、あるいは開始直後の不安定な時期であり、損傷に対して非常にデリケートである。また、チェックポイント機構が十分に働かない場合、損傷を抱えたまま分裂に進んでしまうリスクも高い。
- 後期S期(高抵抗性)の理由: DNAを複製中、あるいは完了直後であるため、すぐ横にテンプレートとなる「もう一本のDNA(姉妹染色分体)」が存在する。これにより、「相同組み換え修復」という極めて正確な修復が可能になり、ダメージを帳消しにできるため。
2-3. 分裂遅延と「G2ディレイ」
細胞は放射線を受けると、無理に分裂を進めようとせず、一旦立ち止まる。これを分裂遅延と呼ぶ。
- G2期の延長:放射線が当たると、特に「G2期」が長くなる(G2ブロック)。M期に入る前に必死にDNAの傷を直そうとする「点検・修理の時間」を確保するためである。
- 遅延の目安:照射線量に比例して長くなり、10Gyまでは1Gyあたり約1時間遅れる。
2-4. 細胞の安全装置「チェックポイント」
細胞周期には、次のステージに進んで良いかを厳格に審査する警備システム(チェックポイント)が存在する。
- p53(がん抑制遺伝子): DNAに傷を見つけると細胞周期を一時停止させ、修理を命じる。修理不能ならアポトーシスを命じる「現場監督」のような存在。
- ATM遺伝子: DNAの2本鎖切断を検知するセンサーの役割。このATM遺伝子が欠損していると、チェックポイントが機能せず、傷を抱えたまま分裂してしまう(毛細血管拡張性運動失調症)。このため放射線感受性が極めて高く、高確率でがんになる。
第3章:DNAの構造と損傷(分子レベルの破壊)
放射線が細胞を殺す際の「最大の標的」はDNAである。放射線生物学において、DNAがどのような構造をしており、放射線によって「どう壊されるのか」を理解することは、すべての基礎となる。
3-1. DNAの基本構造と「壊れやすい場所」
DNAは、遺伝情報を記録するための巨大な分子(生体高分子)である。まずは国家試験で問われる基本パーツとその結合の性質を整理する。
- DNAの基本骨格:2本の「ポリヌクレオチド」の鎖が絡み合った二重らせん構造である。
- ヌクレオチドの構成:塩基 + 糖(デオキシリボース) + リン酸 の3点セット。
- 縦の繋がり(強固):糖とリン酸は「ホスホジエステル結合」という非常に強い結合で繋がっており、DNAの「背骨」を形成している。
- 横の繋がり(脆弱):左右の塩基同士は「水素結合」という比較的弱い結合で対を作っている。
- 塩基のペアの絶対法則(A-T、G-C):
- プリン塩基:アデニン(A)、グアニン(G)
- ピリミジン塩基:チミン(T)、シトシン(C)
- DNA複製の材料:細胞がDNAをコピーする際に使われる部品は、dATP、dTTP、dGTP、dCTP(デオキシリボヌクレオチド三リン酸)と呼ばれる。
3-2. DNA損傷の法則:「頻度」と「致死率」の逆転現象
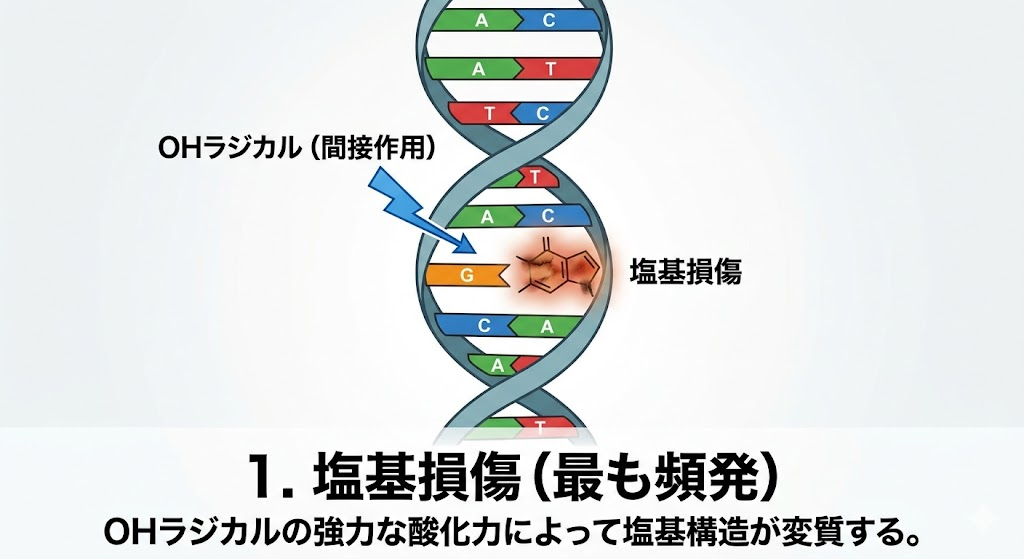
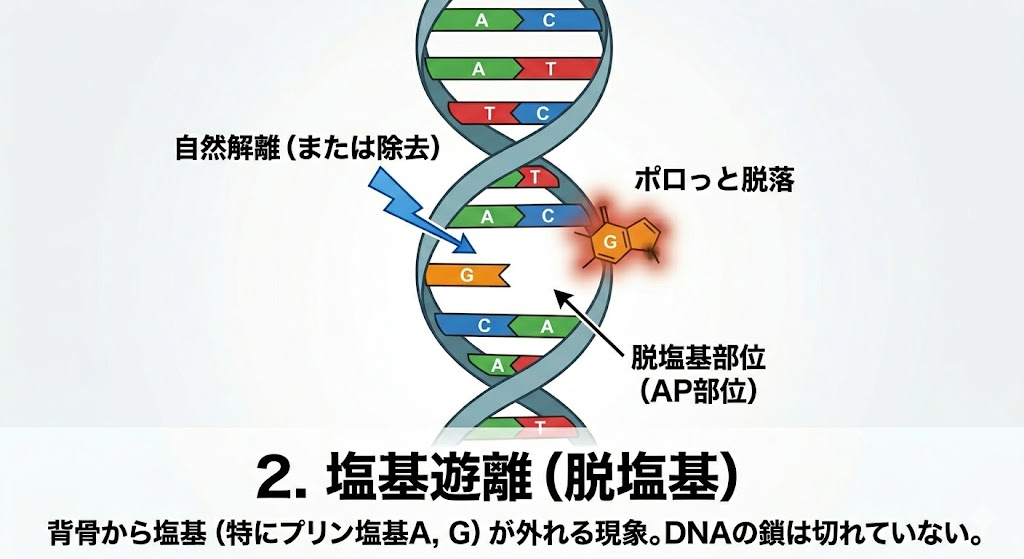
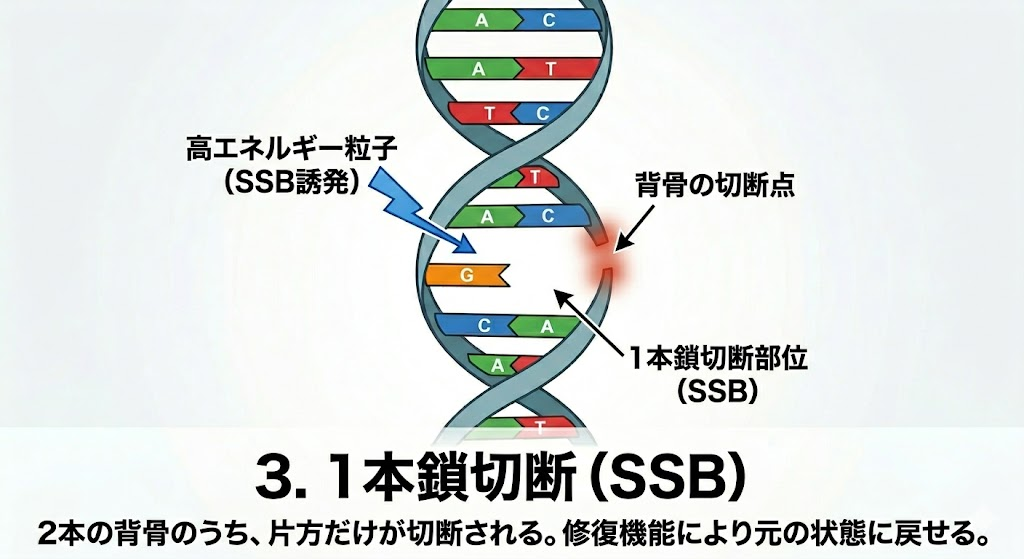
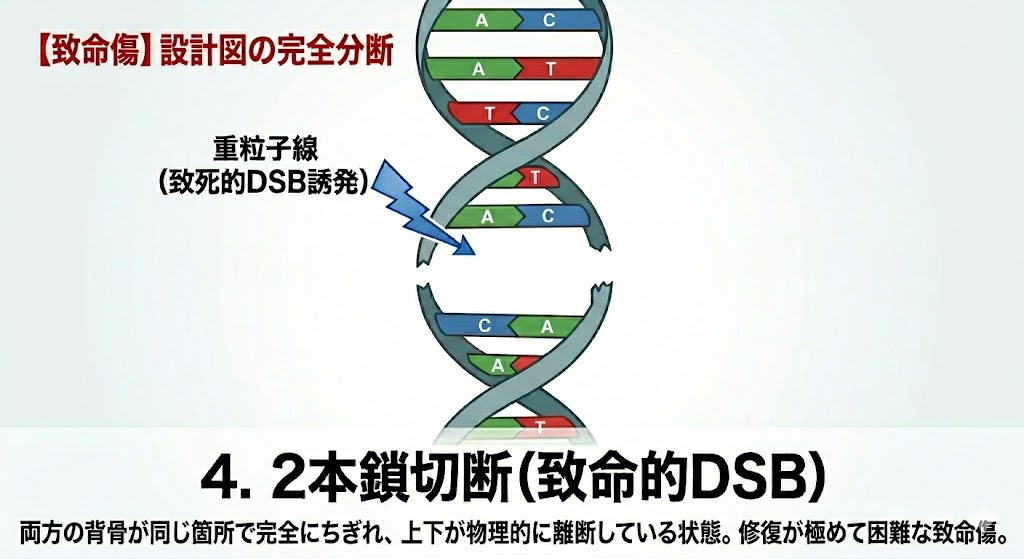
放射線がDNAを攻撃した際、複数の種類のダメージが発生する。ここで絶対に覚えるべき国試の鉄則は、**「起きやすい損傷ほど細胞にとっては大した傷ではなく、起きにくい損傷ほど致命傷になる」**という逆転の法則である。
【発生頻度が高い(=細胞死への影響は少ない)順】
- 塩基損傷:最も頻発する。第2章で学んだ水からの「OHラジカル(間接作用)」の強力な酸化力によって、塩基の構造そのものが変質してしまう現象。
- 塩基遊離(脱塩基):塩基がDNAの背骨からポロっと取れて抜け落ちてしまう現象。分子の構造上、**プリン塩基(A、G)**に起こりやすい。
- 1本鎖切断(SSB:Single Strand Break):2本あるDNAの背骨(ホスホジエステル結合)の片方だけが切断される。もう片方の鎖が無傷で残っているため、後述する修復機能によって容易に元の状態に戻せる。
- 2本鎖切断(DSB:Double Strand Break):最も発生頻度は低いが、細胞死への影響が最も大きい致命傷。 向かい合った2本の鎖が、数塩基対以内の近距離で同時に切断される現象。設計図が完全に分断されるため、修復が極めて困難になる。
3-3. 特殊な損傷:紫外線による「ピリミジンダイマー」
国家試験で引っかけとして頻出するのが、放射線(X線やγ線)ではなく**「紫外線(非電離放射線)」**によって生じる特有の損傷である。
- ピリミジンダイマー(ピリミジン二量体): 紫外線エネルギーによって、同じ鎖の上で隣り合っているピリミジン塩基同士(特にTとT(チミンダイマー))が、異常な共有結合を作ってくっついてしまう現象。DNAのらせん構造が歪み、複製や読み取りがストップしてしまう。
3-4. 高LET放射線の脅威「クラスター損傷」
X線などの低LET放射線は、エネルギーの落とし方が「まばら」であるため、DNA上にポツン、ポツンと単独の損傷(1本鎖切断や塩基損傷)を作るにとどまる。
しかし、重粒子線やα線などの高LET放射線は、エネルギーを極めて「密集」して落とす。そのため、DNAの局所的(1〜2巻きの狭い範囲内)に、2本鎖切断、1本鎖切断、塩基損傷などが同時に複数個所集中して発生する。 これを**局所多重損傷(クラスター損傷)**と呼ぶ。クラスター損傷は細胞の修復システムを完全にキャパオーバーさせるため、高LET放射線のRBE(生物学的効果比)が高くなる最大の物理的根拠となっている。
第4章:DNA修復メカニズム(細胞の決死のリカバリー)
放射線によってDNAが破壊されても、細胞はただ死を待つわけではない。即座に修復システムが稼働する。傷の深さ(1本鎖か、2本鎖か)によって、修復にかかる時間と「正確性」は全く異なる。
4-1. 1本鎖損傷の修復(迅速かつ正確)
1本鎖切断(SSB)や塩基損傷の場合、向かい側の無傷な鎖が「設計図」としてそのまま残っている。そのため、大部分が数分以内という短時間で、元の状態に正確に修復される。
- 光回復:光回復酵素を用いて、損傷(主にピリミジンダイマー)を直接消去し元に戻す。
- 除去修復:傷ついた塩基やヌクレオチド、あるいはミスマッチ(間違った塩基対)をハサミで切り取り、新しく正しいパーツをはめ込んで糊付けする修復。
- 組み換え修復:複製時に損傷を一旦スキップし、後から正常な鎖の情報を借りて穴埋めをする修復。
【色素性乾皮症(XP)】 遺伝疾患に**「色素性乾皮症」がある。
これは、紫外線によってできた傷(ピリミジンダイマー)を直すための「ヌクレオチド除去修復」の機能が先天的に欠損している**病気である。
そのため、紫外線に対して極めて高い感受性を示し、高確率で皮膚がんを発症する。
4-2. 2本鎖切断(DSB)の修復:究極の2択
両方の鎖が同時に千切れる「2本鎖切断」は、設計図を完全に失った状態である。修復には数時間の時間を要し、ミスが起きれば突然変異や細胞死(アポトーシス)に直結する。
細胞は、この致命傷に対して2つの全く異なるアプローチで修復を試みる。
① 非相同末端結合(NHEJ:Non-Homologous End Joining)
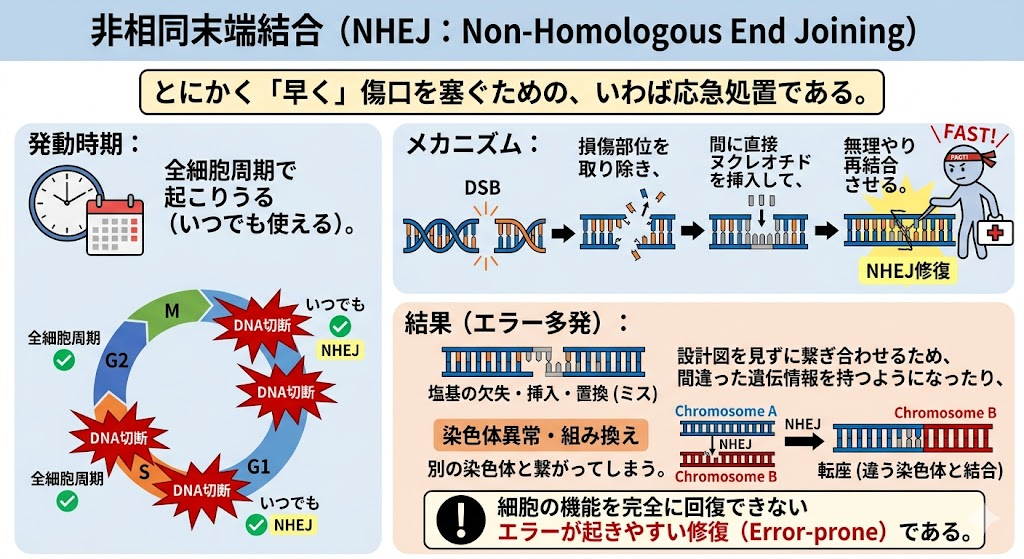
とにかく「早く」傷口を塞ぐための、いわば応急処置である。
- 発動時期:全細胞周期で起こりうる(いつでも使える)。
- メカニズム:切断された端と端の損傷部位を取り除き、間に直接ヌクレオチドを挿入して、無理やり再結合させる。
- 結果(エラー多発):設計図を見ずに繋ぎ合わせるため、間違った遺伝情報を持つようになったり、別の染色体と繋がってしまう(染色体異常・組み換え)ことが多い。細胞の機能を完全に回復できない**エラーが起きやすい修復(Error-prone)**である。
② 相同組み換え修復(HR:Homologous Recombination)
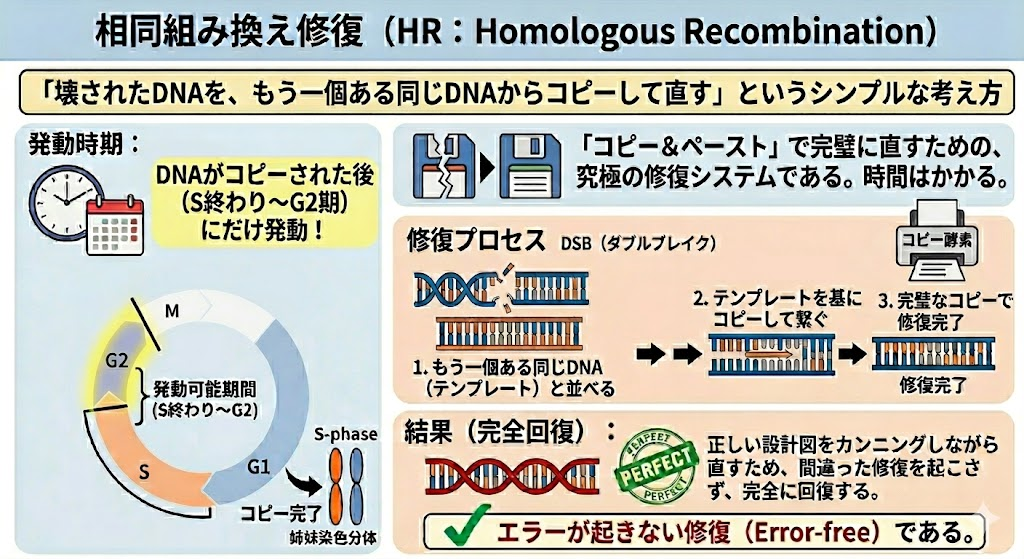
時間をかけてでも「完璧」に直すための、究極の修復システムである。
- 発動時期:S期の終わり〜G2期にしか起こらない。(※ここが超重要!)
- メカニズム:DNAの複製が終わった後、すぐ横には「全く同じ情報を持つ無傷のDNA(姉妹染色分体)」が存在する。ヌクレアーゼやヘリカーゼといった酵素の働きによって、この無傷の姉妹染色分体と交差させ、正常な遺伝情報をテンプレートとして読み込みながらDNAを合成・修復する。
- 結果(完全回復):正しい設計図をカンニングしながら直すため、間違った修復を起こさず、完全に回復する。**エラーが起きない修復(Error-free)**である。
4-3. 伏線回収:なぜ「後期S期」は放射線に最も強いのか?
第2章で「細胞周期の中で、後期S期が最も放射線に強い(抵抗性が高い)」と解説した。その最大の理由は、この**「相同組み換え修復(HR)」がフル稼働できる唯一の時期だから**である。
M期やG1期では、まだDNAが複製されていないため「姉妹染色分体(身代わりの設計図)」が存在しない。そのため、エラーだらけの非相同末端結合(NHEJ)に頼るしかなく、結果として細胞が死にやすい(感受性が高い)のである。
\ あわせて読みたい国試過去問演習 /
今回勉強した基礎知識が、実際の国家試験でどう出題されているかチャレンジしてみよう!
現在、noteにて最新の「第77回 国家試験(午前)」の徹底図解解説を【完全無料】で丸ごと公開中。スマホ対応なのでスキマ時間の復習にも最適!

🎁 効率重視で合格するための「国試対策ロードマップ」を公開中!
当サイトでは、週5〜6でバイトをしながら、ゴルフや旅行も全力で楽しみつつ、夜間学校からストレートで国試160点を叩き出した現役技師の「要領重視の勉強ノウハウ」を1つのマップにまとめています。
「教科書が難しすぎて進まない」「無駄な丸暗記で時間を無駄にしたくない」という方は、まず以下のトップページから、合格までの最短ルートを確認してください!

コメント